
II. Kondice kořisti a biodiverzita – jsou velké šelmy přínosem pro druhovou rozmanitost?
Ve Světě myslivosti č. 12/2009 jsme se věnovali vlivu vlků a rysů na početnost jejich kořisti, což je téma, které patří v souvislosti se šelmami zřejmě k nejdiskutovanějším. Výskyt velkých šelem má však ve vztahu k jejich kořisti také kvalitativní efekty a přinejmenším zajímavá je interakce s dalšími složkami ekosystému - vegetací, rozkladači, níže postavenými predátory nebo vztah mezi různými druhy vrcholových predátorů.
Vliv na spárkatou zvěř
Z mysliveckého hlediska mají význam pro obhospodařování spárkaté zvěře především vlci a rysi. Pro obě šelmy není lov jednoduchou záležitostí - úspěšnost závisí na početnosti kořisti, ročním období a také na tom, zda loví na jimi trvale obývaném území. Dlouholeté průzkumy ukázaly, že jen 20-80 % útoků rysa a 10-49 % útoků vlka končí úspěchem [1,2]. Proto se nejčastější kořistí šelem stává zvěř slabá, mladá (nebo naopak příliš stará), nemocná či hůře smyslově vybavená. Přitom odstranění kondičně podprůměrných jedinců z populace je základním předpokladem úspěšného chovu a v případě srnčí zvěře klíčem ke kvalitním trofejím [3].
Jak se této úlohy zhostil v ČR rys, popsali ve Světě myslivosti č. 3/2003 doc. Koubek a doc. Červený [3]. Příkladem jim byl vývoj průměrné hmotnosti vyvržených srnců v Pošumaví (oblasti se stálým výskytem rysa) a na jižní Moravě (bez trvalého výskytu rysa). Zatímco v letech 1970-2000 průměrná hmotnost srnce na jižní Moravě trvale klesala, v Pošumaví se zvyšovala. Autoři nabízejí dvě možnosti, jak zjištěný nárůst hmotnosti srnčí zvěře na Šumavě vysvětlit: bezchybným průběrným odstřelem na Šumavě a zároveň jeho naprostým selháním na jihu Moravy, nebo instinktivním lovem kondičně špatných jedinců rysem. Přímá souvislost mezi výskytem rysa a vysokou trofejovou kvalitou srnčí není patrná, podrobná analýza zatím nebyla na celorepublikové úrovni provedena [4].
V polských Karpatech by zjištěno, že 82 % laní ulovených v zimě rysem bylo ve velmi špatné tělesné kondici, což byl podíl výrazně odlišný od náhodného vzorku získaného odstřelem [5]. Tatáž studie ovšem selektivní lov v případě vlků neprokázala, což naznačuje, že vlci neloví výhradně nejslabší kořist a za nepříznivých klimatických podmínek (např. hluboký sníh, kde mají vlci výraznou výhodu) se rozdíly v individuální zdatnosti zvěře stírají a kořist vlků pak představuje náhodný vzorek z populace [5]. Přesnější výsledky z polských Karpat přinesl o 20 let později důkladnějším monitoringem Śmietana [6], který celoročně sledoval kořist strženou vlky. Zjistil, že podíl kolouchů ve špatné tělesné kondici byl mezi kořistí vlků výrazně vyšší, než jaký byl náhodný vzorek v populaci. Bylo také prokázáno, že mladí jeleni byli především v zimě a na jaře častější kořistí vlků než laně, což korespondovalo s jejich menší tukovou zásobou, a tedy i horší tělesnou kondicí ve srovnání s laněmi [6].
Výrazný podíl hendikepovaných jelenů (65 %) a mladých divočáků (80 %) v kořisti vlka byl zjištěn i na Slovensku. V oblastech trvalého výskytu vlků byli navíc jeleni o 10-12 % méně promořeni plicními parazity a z těchto oblastí pocházelo také 80 % zlatých trofejí jelenů a černé zvěře [7]. Zjištěné poznatky nejsou nijak překvapivé a potvrzují slova zakladatele československého mysliveckého výzkumu, prof. Julia Komárka, která citujeme z jeho nejznámější knihy Lovy v Karpatech (1942): „Výběr, který vlci mezi vysokou zvěří odpradávna v Karpatech prováděli, byl vždy lepší než náš tak zvaný výběrný odstřel. (...) Výsledky vlků v pěstování jelenů v Karpatech byly rozhodně lepší, než když se toho ujali myslivci sami a začali puškou provozovat nápravu přírody." [8].
Z výsledků publikovaných studií je zřejmé, že s výjimkou Polských Karpat [6] loví vlci i rysi výrazně častěji samičí zvěř, čímž v mnoha honitbách napravují poměr pohlaví vychýlený ve prospěch samičí zvěře [7, 9, 3].
Nedávná studie provedená na Slovensku objevila nápadnou souvislost mezi absencí vlků a výskytem klasického moru prasat (KMP): pouhých 7 % z případů tohoto závažného onemocnění se vyskytlo v oblastech s trvalým výskytem vlků. Nositeli nákazy jsou v 95 % selata a lončáci do 1,5 roku věku, kteří bývají nejčastější kořistí vlků. Mimo souvislý areál výskytu vlka divočáci nemají přirozeného nepřítele a asanace případů KMP nezabránila šíření nákazy do přilehlých oblastí. V oblastech výskytu vlka se mor do dalších oblastí nešířil [10, 11].

Na Slovensku spadalo do oblasti bez trvalého výskytu vlků (zeleně) v letech 1994-1998 93 % případů onemocnění klasického moru prasat. Body zobrazují jednotlivé případy onemocnění divokých prasat, kroužky ojedinělý výskyt vlka mimo souvislý areál. Upraveno podle Finďa [10] a Strnádové [11].
V několika oblastech evropské části bývalého Sovětského svazu bylo zaznamenáno, že v období silného odstřelu vlků se desetinásobně zvýšil podíl losů hynoucích kvůli nemocem a zároveň se snížila početnost populace losů. Když byl odstřel vlků zastaven, šestinásobně se zvedl podíl zvěře ulovené vlky a početnost losů uhynulých v důsledku onemocnění klesla na minimum [12]. Poslední dva příklady potvrzují, že funkce velkých šelem v ekosystému není jen selektivní, ale také sanitární. Slovenský biolog J. Voskár považuje vlky vzhledem k vysokému predačnímu tlaku na lišky také za hlavní přirozené regulátory vztekliny [7].
Biodiverzita
Návrat velkých šelem představuje sám o sobě obohacení přírody o druhy, které byly v důsledku intenzivního využívaní krajiny a lovu vyhubeny v některých oblastech již před staletími, jinde - především na východní Moravě - před desetiletími. Výskyt přirozených predátorů však má i další efekt. Pod pojmem biodiverzita nemáme na mysli jen rozmanitost biologických druhů, ale také genetickou nebo ekosystémovou diverzitu [13]. Poněkud širší definice chápe biodiverzitu také jako interakci mezi všemi druhy a s tím související ekologické a behaviorální procesy [14, 15]. Nejprve se zaměřím na oblast biodiverzity, která souvisí se snižováním početnosti kopytníků - tedy efektem, jehož může být docíleno i vhodným mysliveckým hospodařením. Následně se budu věnovat přínosům, jichž by bez velkých šelem nešlo většinou dosáhnout.
Početnost kopytníků
Jak bylo popsáno v prvním díle článku (Svět myslivosti č. 12/2009), možnosti velkých šelem aktivně regulovat početnost kořisti jsou v našich podmínkách zřejmě omezené. Lokálně však může být jejich vliv na snížení početnosti spárkaté zvěře významný.
Populace volně žijících kopytníků, které se vymkly kontrole a jejichž stavy se pohybují nad únosnou kapacitou prostředí, závažně poškozují vegetační kryt a snižují druhovou diverzitu. Mnoho studií potvrzuje, že nadměrné spásání ničí přirozenou strukturu biotopů a připravuje o domov např. mnoho druhů ptáků [16]. Těm zároveň ubývají zdroje potravy [17]. Současně ubývají i lesní byliny, včetně chráněných druhů [18]. U nás jsou ohroženy některé přírodní rezervace, které si zachovaly přírodě blízké druhové složení, ale v důsledku intenzivního okusu zvěře zde neprobíhá přirozená obnova dřevin zastoupených v mateřském porostu. Vážné poškození okusem bylo v letech 2001-2002 zjištěno v pěti ze sedmi zkoumaných českých národních přírodních rezervací [19, 20].
V mnoha studiích byl sledován vliv vysoké hustoty zvěře na bezobratlé živočichy. Např. v oplocených plochách ve Skotsku se vyskytovalo čtyřikrát více larev motýlů, třikrát více blanokřídlých a dvakrát více brouků, dvoukřídlých i pošvatek než v okolním, zvěří intenzivně spásaném lese. Ve skotských poměrech tak jelení zvěř významně omezuje dostupnost potravy nejen pro lesní hmyzožravé ptáky, ale i pro tetřeva hlušce, jehož kuřata jsou z velké části odkázána na motýlí larvy, mravenčí kukly a listy borůvky (která navíc dosahuje na plochách spásaných jeleny poloviční výšky ve srovnání s oplocenými plochami) [21]. Nicméně jiné studie ukázaly, že zvýšená intenzita pastvy mnohým, často chráněným druhům prospívá, proto obecný závěr ve vztahu k bezobratlým nelze učinit. Intenzivnější pastva by proto v budoucnu mohla být přínosná v některých nížinných světlinových lesích, které jsou domovem řady chráněných druhů motýlů. V nich by se mohlo uplatňovat výmladkové hospodaření, které je pro množství vzácných druhů hmyzu existenční podmínkou [22].
Změna chování spárkaté zvěře
Populační hustota býložravců není jediným faktorem ovlivňujícím biodiverzitu. Ojedinělá možnost sledovat obnovu ekosystému se naskytla biologům v americkém národním parku Yellowstone. Přestože podmínky, které velkým šelmám poskytuje krajina střední Evropy, se výrazně liší, považuji za užitečné seznámit čtenáře Světa myslivosti se zmíněným výzkumem jako modelem popisujícím širší souvislosti procesu predace.
V Yellowstonském národním parku byli vlci vyhubeni ve dvacátých letech 20. století. V letech následujících po vyhubení vlků jelení zvěř intenzivně spásala údolní nivy řek a obnova listnatých dřevin (vrby, topoly) se zcela zastavila. Ani redukce početních stavů jelení zvěře odstřelem neměla pro obnovu ekosystémů význam.
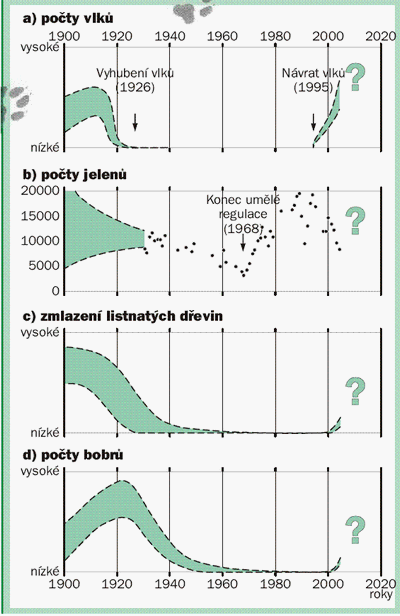
Historické trendy severní části Yellowstonského národního parku (USA) od roku 1990. Šířka zelených pruhů je odrazem nejistoty, s jakou byla na základě literatury rekonstruována početnost živočichů a rostlin. Snížení početnosti jelenů (b) nebylo v období 1996-1998 primárně způsobeno predací vlků, ale tuhými zimami. Nárůst početnosti bobrů v letech 1900-1920 souvisel s obnovou populací po silném loveckém tlaku v 19. století. Další souvislosti jsou vysvětleny v doprovodném textu. Upraveno podle Rippleho a Beschty [23].
V roce 1995 proběhla reintrodukce vlka a pouhých sedm let po jeho návratu do Yelowstone bylo v říčních nivách pozorováno nebývalé zmlazení listnatých dřevin. Nejvýznamnější přínos návratu vlků spočíval ve změně chování kopytníků: jelení zvěř už si nemohla dovolit trávit dlouhý čas na lokalitách, kde jí hrozilo nebezpečí od predátorů a začala častěji měnit stanoviště a využívat jiné, spásáním vegetace méně zranitelné biotopy. S obnovou říčních porostů, klíčových pro diverzitu a funkci celých ekosystémů, se vrátili bobři, kořeny vrb stabilizovaly říční koryta, obnovily se mokřady, zvýšila se početnost vodních bezobratlých, ptáků i další fauny.

Vrbové porosty podél potoka Blacktail Creek v Yellowstonském národním parku (USA) na jaře 1996 (vlevo) a v létě 2002 (vpravo). Na snímku z roku 1996, pořízeném po 70letém období bez přítomnosti vlků, jsou zachyceny silně spásané vrby a jehličnany. 7 let po návratu vlků (2002) je evidentní, že se vrby dokázaly z tlaku zvěře vymanit. Foto: Správa Yellowstonského národního parku a W. Ripple.
Efekt, který znamenal návrat vlků, se tak prostřednictvím tzv. „predačního risku" - obavy kořisti vlků z útoku - kaskádově přenesl na nižší trofické úrovně a ukázal, jak může vrcholový predátor pozitivně ovlivnit ekosystém i bez toho, že by přímo snižoval početnost své kořisti [23].
Podobný efekt lze pozorovat i v našich podmínkách: v oblastech výskytu velkých šelem zvěř mění své návyky, resp. vrací se ke svému přirozenému chování - stává se opatrnější, mění častěji stanoviště a žije skrytějším životem [24, 7]. Z hlediska ochrany přírody jsou nejcennějšími územími zpravidla přírodní rezervace, které jsou kvůli bohaté potravní nabídce atraktivní i pro zvěř. V řadě z nich je v důsledku lokálního přemnožení zvěře a nedostatku úživných biotopů blokována přirozená obnova [19, 20]. Zatím nebyl proveden exaktní výzkum, ale pozorování některých lokalit v Beskydech, kde se rysi vyskytují pravidelně od 70. let [25, 26, 27] a vlci od poloviny 90. let dvacátého století [28], ukazují na úspěšnou obnovu zejména jedle bělokoré, dorůstající i bez umělé ochrany. Zda jde skutečně o důsledek změny chování zvěře v oblíbených lovištích rysa nebo vlka však zatím nevíme.
Kadávery
Velké šelmy mohou zvyšovat biodiverzitu také podporou mrchožroutů a rozkladačů, kteří se přiživují na zbytcích jimi stržené kořisti. V Białowiezském národním parku bylo zjištěno, že zbytky kopytníků jsou důležitým potravním zdrojem pro 30 druhů malých a středně velkých savců a ptáků. V případě absence vrcholových predátorů jsou kadávery přístupné jen sezónně, nejčastěji v zimě. Přítomnost vlka a rysa tak představuje celoroční přísun živočišných zbytků [29].
Výzkumu kadáverů jako zbytků po kořisti velkých predátorů byl věnován také zvláštní workshop na loňské evropské konferenci ochrany přírody konané v Praze. Chris Wilmers z Kalifornské univerzity na příkladu Yellowstonského národního parku poukázal na fakt, že s přicházejícím globálním oteplováním a menší sněhovou pokrývkou lze v zimě očekávat i menší množství kadáverů. Celoroční přísun kadáverů jako zbytků po kořisti vlků přitom umožňuje přežít také dalším živočichům závislým na tomto typu potravy a tlumit tak jeden z důsledků globálního oteplování na místní ekosystémy. John Linnell z Norského institutu pro výzkum přírody však konstatoval, že v norských podmínkách je množství zbytků srnčí zvěře (kořist rysa) relativně malé (0,14 kg / hektar / rok) a vzhledem k jiným potravním zdrojům zřejmě není v tomto smyslu úloha rysa zásadní.
Rys versus tetřev
Výskyt velkých šelem často vyvolává otázku, zda tito predátoři nemohou ohrožovat další chráněné živočichy, například tetřevovité ptáky (tetřev hlušec, tetřívek obecný, jeřábek lesní).
Je pravda, že rys se živí i menšími savci a ptáky, pro tetřevy však rozhodně není hlavní hrozbou. Na Šumavě byly pozůstatky tetřevovitých (jeřábka lesního a tetřeva hlušce) nalezeny jen v 3,6 % vzorků trusu [26]. Mnohem významnějšími predátory kurovitých jsou ve skutečnosti menší šelmy - liška a kuna. M. Saniga ve slovenských Karpatech zjistil, že ze 49 zničených tetřevích hnízd vybrala 22 % liška, kuna nebo lasice, 9 % prase divoké a 3 % medvěd. Z ptačích predátorů měli význam krkavci a sojky, kteří byli odpovědní za 18 % zničených hnízd [30].
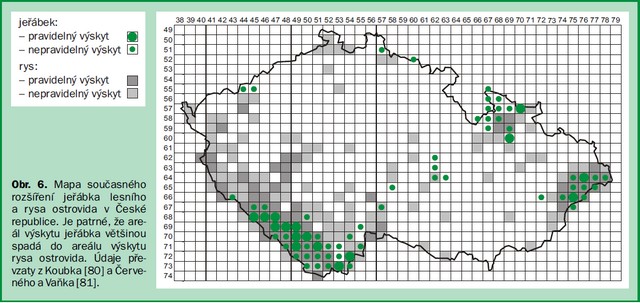
Z odchovaných tetřevů vypouštěných na Šumavu, do Českého lesa a Krušných hor způsobila 76 % všech známých úhynů (telemetricky sledovaných jedinců) liška nebo kuna [31]. Důkazem, že jak tetřev, tak jeřábek mohou koexistovat s velkými šelmami na stejném území, může být společný výskyt této zvěře v celém horském pásmu Karpat nebo překryv areálů jeřábka lesního a rysa ostrovida v České republice (viz obr. 6). Podíváme-li se blíže na konkrétní lokality, zjistíme, že například horský masiv Smrk v Beskydech s pravidelným výskytem všech velkých šelem [32] je zároveň jednou z nejčastějších oblastí výskytu tetřevovitých ptáků [33].
Ve skutečnosti jsou tedy vlci a rysi spíše prospěšní tetřevovitým tím, že redukují početnost predátorů (lišek nebo divočáků), než že by je sami ohrožovali predací. Pro udržení populací tetřeva a jeřábka je však potřebné provádět současně komplex opatření, zahrnující šetrné lesnické zásahy, které zvyšují potravní nabídku a členitost podrostu, regulaci rekreačně-sportovních aktivit a turistického ruchu a snížení počtu malých šelem, černé a jelení zvěře. [33].
Regulace malých šelem
Studiem dlouhých časových řad ve Švédsku od roku 1828-1910 bylo zjištěno, že narůstající početnost malých predátorů (lišek) způsobují dva faktory: 1. změna využití zemědělské půdy, jež vedla ke zvýšení produktivity prostředí včetně nárůstu početnosti kořisti malých šelem (především hlodavců) a 2. potlačením vrcholových predátorů (vlk, rys), kteří mohou početnost malých šelem regulovat „shora" [34]. Fakt, že liška se může stát potravou rysa ostrovida, známe i ze Šumavy, kde byla liščí srst nalezena v 5 % vzorků [26].
Důkladnější telemetrická studie ve Švédsku prokázala, že rys se může významně podílet na redukci lišek: 50 % známých případů úmrtí způsobil rys. Početnost lišek ve sledovaném regionu poté, co území znovuosídlil rys, během 10 let trvale klesala. Následné analýzy vyhodnotily predaci rysem jako nejpravděpodobnější příčinu tohoto poklesu [35]. Rovněž současné zkušenosti některých myslivců v Beskydech naznačují, že početnost lišek v oblasti s trvalým výskytem rysa je menší [36]. Na Slovensku byl zjištěn výrazný predační efekt vlka na populaci lišek a toulavých psů [7], podobně řada amerických studií prokázala nárůst početnosti jiných menších šelem - kojotů v oblastech, kde byli vlci vyhubeni, nebo naopak jejich pokles, pokud se vlci vrátili [2].
Regulace bobrů
Mnoho druhů středně velkých šelem občas uloví bobra, ale pravidelně tak činí pouze vlk, pro kterého bobr může znamenat jeden z nejdůležitějších druhů alternativní kořisti. Ačkoliv to v našich podmínkách není prozatím aktuální, je třeba zmínit, že v řadě zemí, například v Estonsku, se vlci podílejí na regulaci bobří populace [37].
Vztahy mezi vrcholovými predátory
Pozorování přímého kontaktu mezi vlky, rysy a medvědy jsou v evropských podmínkách poměrně vzácná. Vzhledem k převažující rostlinné stravě medvěda a jeho výrazné preferenci konzumace kadáverů před aktivním lovem není konkurence s vlkem a rysem příliš pravděpodobná. Díky výbornému čichu ovšem může medvěd nacházet a využívat kořist, strženou vlkem nebo rysem.
Někdy se uvádí, že přítomnost vlků negativně ovlivňuje výskyt rysů (případně naopak). Ve skutečnosti však pro podobné úvahy není zatím odborné zdůvodnění. Vlk obecný a rys ostrovid se vyskytují na významné většině svého euroasijského areálu od východní Evropy po Dálný východ sympatricky (tedy společně). Detailní telemetrické sledování vlků a rysů v Bialowiežském národním parku v 90. letech a následná analýza všech dat však ukázala, že domovské okrsky rysů a teritoria vlků se významně překrývaly (76 % vlčích teritorií a 50% rysích domovských okrsků) a bylo také vícekrát zaznamenáno přiblížení obou druhů na vzdálenost až 250 metrů. Polští vědci nenašli ani negativní, ani pozitivní vazbu mezi výskytem vlků a rysů, oba druhy šelem využívaly prostor nezávisle. Důvodem je pravděpodobně specializace obou šelem na různé druhy kopytníků (rysa na srnčí a vlka na jelení zvěř) a různorodost a členitost biotopů (rysům vyhovuje členitější terén) [38].
Budoucnost ?
Sociální vyspělost velkých šelem a vztahy mezi nimi a dalšími složkami ekosystémů jsou často poměrně složité a souvislosti široké. Tolik ceněné vlastnosti zvěře jsou výsledkem dlouhé evoluce mezi predátorem a kořistí.
Přizpůsobivost velkých šelem je také poměrně velká a klišé, že vlci a rysi jsou symbolem divočiny neplatí: například vlci se z několika smeček ve východním Německu (Lužici) šíří k Berlínu nebo přes Polsko k běloruským hranicím. Oproti našim západním sousedům se náš stát zdá být vhodným prostředím a je otázka, proč se např. vlci nebo rysi trvale neusadili v našich pohraničních pohořích. Velké šelmy alespoň ukázaly, že s námi dovedou žít. Závisí na nás, zda jsme ochotni i my se s nimi bez předsudků podělit o krajinu.
Mgr. Miroslav Kutal
Literatura:
[1] Hell P., Slamečka J. & Gašpárík J., 2004: Rys a divá mačka v slovenských Karpatoch a vo svete. PaRPRESS Bratislava. 162 pp.
[2] Mech D. & Boitiani L. (eds.), 2003: Wolves: behavoiur, ecology and conservation. The University of Chicago Press, Chicago, 448 pp.
[3] Koubek P) : 8-10.
[4] Engan J. H. (in prep.) Roe deer trophies in Czech Republic on maps - Srnčí trofeje v ČR v mapovém vyjádření - Lysá nad Labem 2005 (nepublikovaná studie).
[5] Okarma H., 1984: The physical condition of red deer falling a prey to the wolf and lynx and harvested in the Carpathian Mountains. Acta Theriologica 29 (11-25): 283-290.
[6] Śmietana W., 2005: Selectivity of wolf predation on red deer in the Bieszczady Mountains, Poland. Acta Theriologica 50 (2): 277-288.
[7] Voskár J., 1993: Ekológia vlka obyčejného (Canis lupus) a jeho podiel na formování a stablite karpatských ekosystémov na Slovensku. Ochrana prírody 12: 241-276.
[8] Komárek J., 1942: Lovy v Karpatech. Státní zemědělské nakladatelství, Praha, 5. vydání, 285 pp.
[9] Červený J., 2006: Myslivec a rys, dva lovci a jedna kořist - srnčí zvěř. Svět myslivosti 7 (3): 8-11.
[10] Finďo S., 2002: Potravná ekológia vlka (Canis lupus) v Slovenských Karpatoch. Výskum a ochrana cicavcov na Slovensku V, Zborník referátov z konferencie Zvolen.Bánská Bystrica: 43-55.
[11] Strnádová J., 2000: Predačný efekt vlka dravého (Canis lupus L.) na populáciu diviačej zveri a jeho význam v dynamike klasického moru ošípaných diviakov na Slovensku. Diplomová práce. Přírodovědecká Fakulta Univerzity Komenského v Bratislavě, 55 pp.
[12] Filonov C., 1980: Predator-prey problems in nature reserves of the European part of the RSFSR. Journal of Wildlife Management 44 (2): 389-396.
[13] Primack R. B., Kindlman P. & Jersáková J., 2001: Biologické principy ochrany přírody. Portál, Praha, 352 pp.
[14] Linnell J. D. C., Promberger C., Boitani L., Swenson J. E., Breitenmoser U. & Andersen R., 2005: The linkage conservation strategies for large carnivores and biodiverzity: the view from the „half full" forests of Europe: 381-399. In: Ray C., Redford K. H., Steneck R. S. & Berger J., 2005: Large carnivores and the conservation of biodiverzity. Island Press, Washington D.C., USA, 526 pp.
[15] Pyare S. & Berger J., 2003: Beyond demography and delisting: ecological recovery for Yellowstone's grizzly bears and wolves. Biological Conservation 113: 63-73.
[16] Gill R. M. A. & Fuller R. J., 2007: The effects of deer browning on woodland structure and songbirds in lowland Britain. Ibis 149 (Supp. 2): 119-127.
[17] Allombert S., Stockton S. & Martin J., 2005: A natural experiment on the impact of overabundant deer on forest invertebrates,. Conservation Biology 19 (6): 1917-1929.
[18] Míchal I., 1992: Obnova ekologické stability lesů. Academia, Praha, 169 pp.
[19] Čermák P. & Mrkva R, 2003: Browsing damage to broadleaves in some national nature reserves (Czech Republic) in 2000-2001, Ekológia (Bratislava) 22 (3): 132-141.
[20] Čermák P. & Mrkva R., 2003: Vliv mysliveckého hospodaření na vývoj dřevinné vegetace. Lesnická práce 82 (6).
[21] Petříček V. & Míchal I. (eds.), 2002: Péče o chráněná území: 2. Lesní společenstva. Agentura ochrany přírody a krajiny ČR, Praha, 713 pp.
[22] Konvička M., Čížek L. & Beneš J., 2004: Ohrožený hmyz nížinných lesů: ochrana a management. Sagittaria, Olomouc, 79 pp.
[23] Ripple W. J. & Beschta R. L., 2004: Wolves and the ecology of fear: can predation risk structure ecosystems? BioScience 54 (8): 755-766.
[24] Červený J., 2006: Myslivec a rys, dva lovci a jedna kořist - srnčí zvěř. Svět myslivosti 7 (3): 8-11.
[25] Kunc L., 1996: Lynx (Lynx lynx) in the Moravskoslezské Beskydy Mts. Acta Sc. Nat. Brno, 30 (3): 58-63.
[26] Červený, J., Koubek, P. & Bufka, L., 2006: Velké šelmy v České Republice. IV. Rys ostrovid. Vesmír 85 (2): 86-94.
[27] Kunc L., 1999: Můj přítel rys. Víkend, 110 pp.
[28] Bartošová D., 1998: Osud vlků v Beskydech je nejistý. Veronica, 11 (1): 1-7.
[29] Jędrzejewski W. & Jędrzejewska B., 2005: Large carnivores and ungulates in European temperate forest ecosystems: bottom up and top down control: 230-246. In: Ray C., Redford K. H., Steneck R. S. & Berger J., 2005: Large carnivores and the conservation of biodiverzity. Island Press, Washington D.C., USA, 526 pp.
[30] Saniga M. 2002: Nest loss and chick mortality in capercaillie (Tetrao urogallus) and hazel grouse (Bonasa bonasia) in West Carpathians. Folia Zool. 51(3): 205-214.
[31] Marhoul P. & Volf O., 2005: Hodnocení realizace prvních pěti let záchranného programu tetřeva hlušce v České republice. Agentura ochrany přírody a krajiny ČR, Praha, 30 pp + VI příl.
[32] Telnarová B., 2004: Výskyt velkých šelem v prostoru horského masivu Smrk. Ostravská univerzita v Ostravě, Přírodovědecká fakulta, Katedra biologie a ekologie, Ostrava, 54 pp. + 28 příl.
[33] Jůzová B. & Krupa M: 2006 Přírodě blízké hospodaření v lesích s ohledem na zachování ptačích druhů (tetřev hlušec a jeřábek lesní) jako předmětů ochrany v Ptačí oblasti Beskydy. Nepublikovaná studie Agentury ochrany přírody a krajiny ČR, 47 pp. + příl.
[34] Elmhagen, B. & Rushton S. P., 2007: Trophic control of mesopredators in terrestrial ecosystems: top-down or bottom-up? Ecology Letters 10: 197-206.
[35] Helldin J. O., Liberg O. & Glöersen G., 2006: Lynx (Lynx lynx) killing red foxes (Vulpes vulpes) in boreal Sweden - frequency and population effects. Journal of Zoology. Journal of Zoology 270 (4): 657-663(7).
[36] Sobotka R., 2007: Pytláci v Beskydech. Víkend, Líbezníce, 158 pp.
[37] Lõhmus A., 2001 (ed.): Status of Large Carnivore Conservation in the Baltic States: Large Carnivore Control and Management Plan for Estonia, 2002-2011. Council of Europe, Strassburg, 53 pp.
[38] Schmidt K, Jędrzejewski W, Okarma H.& Kowalczyk R, 2009: Spatial interactions between grey wolves and Eurasian lynx in Białowieża Primeval Forest, Poland. Ecological Research 24 (1): 1440-1703.
Předchozí článek: I. Početnost - jak moc regulují šelmy svou hlavní kořist?

